

Overview
Tail-anchored membrane proteins, which include a number of essential proteins such as SNAREs, contain a single transmembrane domain at their C-terminus requiring post-translational targeting and delivery. Since the identification of the Guided Entry of Tail-anchored proteins (GET) pathway in yeast in 2007, the lab has worked to structurally characterize all proteins involved in targeting and delivery to the ER. Our research into tail-anchored protein targeting has led us to study the pathway in yeast, the protozoan Giardia intestinalis, and humans– resulting in a variety of NMR, x-ray crystal, and cryo-EM structures.
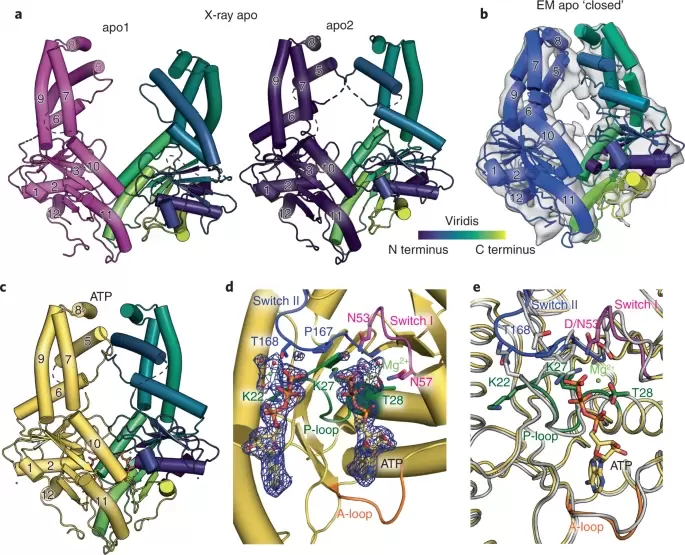
Fry et al. (2022)
|
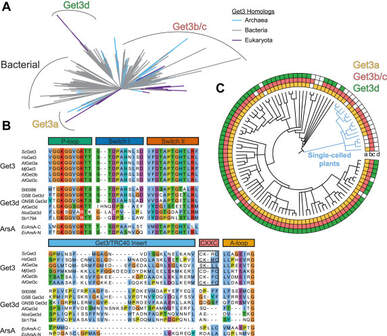
Barlow et al. (2023)
|
See more:
Alexandra N. Barlow*, M. S. Manu*, Shyam M. Saladi, Paul T. Tarr, Yashpal Yadav, Aye M. M. Thinn, Yun Zhu, Arthur D. Laganowsky, William M. Clemons, Jr., Sureshkumar Ramasamy, “Structures of Get3d reveal a distinct architecture associated with the emergence of photosynthesis” Journal of Biological Chemistry (2023)
Michelle Y. Fry, Vladimíra Najdrová, Ailiena O. Maggiolo, Shyam M. Saladi, Pavel Doležal, William M. Clemons, Jr., “Structurally derived universal mechanism for the catalytic cycle of the tail-anchored targeting factor Get3” Nature Structural & Molecular Biology (2022)
Christian Suloway, Justin Chartron, Ma’ayan Zaslaver & William Clemons, Jr. “Model for eukaryotic tail-anchored protein binding based on the structure of Get3.” Proceedings of the National Academy of Sciences (2009) v106 n35 pp14849-54.
Justin Chartron, David Vandervelde, William Clemons, Jr. “Structures of the Sgt2/SGTA dimerization domain with the Get5/UBL4A UBL domain reveal an interaction that forms a conserved dynamic interface.” Cell Reports (2012) v2 n6 pp1620-32.
Harry Gristick, Meera Rao, Justin Chartron, Michael Rome, Shu-ou Shan & William Clemons, Jr. “The structure of a tail-anchor membrane protein-binding complex reveals the regulation of Get3 by Get4.” (2014) Nature Structure & Molecular Biology v21 n5 pp437-42.
Jee-Young Mock, Justin Chartron, Ma’ayan Zaslaver, Yue Xu, Yihong Ye & William Clemons, Jr. “The Bag6 complex contains a minimal tail-anchor targeting module and a 'mock' BAG domain” (2015) Proceedings of the National Academy of Sciences, USA v112 n1 pp106-11.
Alexandra N. Barlow*, M. S. Manu*, Shyam M. Saladi, Paul T. Tarr, Yashpal Yadav, Aye M. M. Thinn, Yun Zhu, Arthur D. Laganowsky, William M. Clemons, Jr., Sureshkumar Ramasamy, “Structures of Get3d reveal a distinct architecture associated with the emergence of photosynthesis” Journal of Biological Chemistry (2023)
Michelle Y. Fry, Vladimíra Najdrová, Ailiena O. Maggiolo, Shyam M. Saladi, Pavel Doležal, William M. Clemons, Jr., “Structurally derived universal mechanism for the catalytic cycle of the tail-anchored targeting factor Get3” Nature Structural & Molecular Biology (2022)
Christian Suloway, Justin Chartron, Ma’ayan Zaslaver & William Clemons, Jr. “Model for eukaryotic tail-anchored protein binding based on the structure of Get3.” Proceedings of the National Academy of Sciences (2009) v106 n35 pp14849-54.
Justin Chartron, David Vandervelde, William Clemons, Jr. “Structures of the Sgt2/SGTA dimerization domain with the Get5/UBL4A UBL domain reveal an interaction that forms a conserved dynamic interface.” Cell Reports (2012) v2 n6 pp1620-32.
Harry Gristick, Meera Rao, Justin Chartron, Michael Rome, Shu-ou Shan & William Clemons, Jr. “The structure of a tail-anchor membrane protein-binding complex reveals the regulation of Get3 by Get4.” (2014) Nature Structure & Molecular Biology v21 n5 pp437-42.
Jee-Young Mock, Justin Chartron, Ma’ayan Zaslaver, Yue Xu, Yihong Ye & William Clemons, Jr. “The Bag6 complex contains a minimal tail-anchor targeting module and a 'mock' BAG domain” (2015) Proceedings of the National Academy of Sciences, USA v112 n1 pp106-11.
The peptidoglycan (PG) is a protective layer conserved in most bacteria, and several enzymes involved in the peptidoglycan biosynthesis have been attractive targets for designing antibacterial agents for decades. A focus of my group has been an interest in chemical reactions that involve glycans at the membrane. This specifically involves two different bacterial systems including peptidoglycan biosynthesis and N-linked glycosylation. For the former, we have currently published studies on a phage lysis protein E that inhibits the central membrane protein MraY demonstrating the sequence requirements, and recently, using single particle electron cryo-microscopy, we demonstrate that protein E bridges two bacterial proteins to form the transmembrane YES complex (MraY, protein E, SlyD).
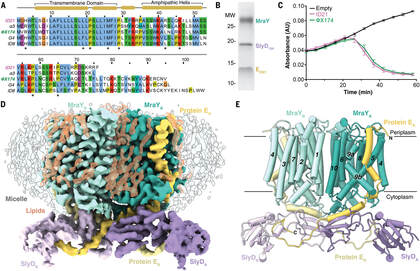
|
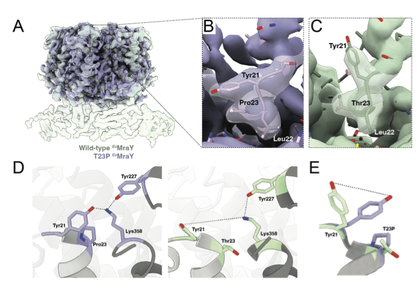
|
|
Orta et al. (2023)
|
Marmont, Orta et al. (2023)
|
See more:
Lindsey S. Marmont, Anna K. Orta, Robin A. Corey, David Sychantha, Ana Fernández Galliano, Yancheng E. Li, Becca W.A. Baileeves, Neil G. Greene, Phillip J. Stansfeld, William M. Clemons Jr, Thomas G. Bernhardt. “A feedback control mechanism governs the synthesis of lipid-linked precursors of the bacterial cell wall” (2023) bioRxiv
https://doi.org/10.1101/2023.08.01.551478.
A. Karen Orta*, Nadia Riera*, Yancheng E. Li, Shiho Tanaka, Hyun Gi Yun, Lada Klaic, William M.Clemons, Jr. “The mechanism of the phage-encoded protein antibiotic from ΦX174.” (2023) Science v381 n6654 http://doi.org/10.1126/science.adg9091
PDBID 8G01 8G02; Free article download link; Perspective by Balbach & Stubbs; Caltech News feature
Hyun Gi Yun, Kyoung-soon Jang, Shiho Tanaka, William M. Clemons, Jr., “The structure of the UDP-Glc/GlcNAc 4-epimerase from the human pathogen Campylobacter jejuni” (2020) bioRxiv https://doi.org/10.1101/2020.09.22.308395 PDBID 7K3P
Lindsey S. Marmont, Anna K. Orta, Robin A. Corey, David Sychantha, Ana Fernández Galliano, Yancheng E. Li, Becca W.A. Baileeves, Neil G. Greene, Phillip J. Stansfeld, William M. Clemons Jr, Thomas G. Bernhardt. “A feedback control mechanism governs the synthesis of lipid-linked precursors of the bacterial cell wall” (2023) bioRxiv
https://doi.org/10.1101/2023.08.01.551478.
A. Karen Orta*, Nadia Riera*, Yancheng E. Li, Shiho Tanaka, Hyun Gi Yun, Lada Klaic, William M.Clemons, Jr. “The mechanism of the phage-encoded protein antibiotic from ΦX174.” (2023) Science v381 n6654 http://doi.org/10.1126/science.adg9091
PDBID 8G01 8G02; Free article download link; Perspective by Balbach & Stubbs; Caltech News feature
Hyun Gi Yun, Kyoung-soon Jang, Shiho Tanaka, William M. Clemons, Jr., “The structure of the UDP-Glc/GlcNAc 4-epimerase from the human pathogen Campylobacter jejuni” (2020) bioRxiv https://doi.org/10.1101/2020.09.22.308395 PDBID 7K3P